同じモンスーン気候の下に自生するCymbidiumとパフィオ。
なぜ、パフィオはバルブを作らなかったのか?
効率の悪い光合成。
オモトにも大葉と・・・天光冠のようなものがあるが、
パフィオのパービセパルム亜属生長の遅さをみると、
オモトの天光冠を連想してしまう。
あまりのスローライフ。
植物の進化にも、逆も真なり・・・・があるようである。
こういう植物と付き合うには、別な理念の生き方が
要求される。
上の写真はウラベニダイモンジソウ。
パフィオパービセパルム亜属の自生地石灰岩と極似
する環境に自生する。
ダイモンジソウは、多湿のところに自生するが、
ウラベニダイモンジソウは、乾燥した石灰岩の
割れ目などでも生息している。
他の植物が生息できない不毛の地。
ダイモンジソウの仲間は、中国にも自生し、
パフィオ パービセパルム亜属と自生地は重なる。
光の集光、反射には共通点があるのかもしれない。
葉の裏が紫のパフィオ
葉の裏が緑色のパフィオ
光の万能型のパフィオ
パフィオの自生地を観察すると、
同じ種であっても弱い光のところにも、
強い光のところにも自生しているので、
どのように栽培したらよいの解からない
ことがある。
光万能型の特性は、光の反射、透過という
相反する機能を持った葉のパフィオである。
過酷な不毛の地に自生するパフィオ。
光の専門型パフィオ。
常に弱い光林の下、霧に深く覆われる場所
に自生するパフィオ。
カランテなどと共通する光である。
太陽高度の算出の仕方
太陽高度=90度ー観測地の緯度+23,4
夏至の時の太陽高度はこの公式で出す。
北回帰線と南回帰線のエリアが熱帯地方である。
北回帰線に近くある都市はホンコン。
南回帰線に近くにある都市はサンパウロ。
ヒマラヤは北回帰線近くから北にあるので、
この地方に自生するランは
四季の変化。
短日長日の変化。
雨期乾期の変化に遭遇することになる。
この複雑な地球のサイクルに合わせて生きる
DNAを持たなければ、いきつづけることは出来ない。
パフィオのパービセパラム亜属などの多くは、
北回帰線以北の自生地であるから、前記の
サイクルの中で生きていることになる。
暑さにも寒さにも対応しなければならない。
日の長さ短さにも対応しなければならない。
多湿、乾燥にも耐えなければならない。
ヒマラヤのモンスーン気候に自生する植物の多くは、
この相反する条件に、何らかの方法で適合しなければ
生きてゆけない。
このような適応性は、熱帯地方の高所に生きるランも
同じである。
低地に自生するランは、この中から、低温に対する
適応性を削除したものである。
例えば、コチョウラン、カトレアなど。
このようなランは栽培は簡単である。少なくとも
生育環境が複雑ではない。寒さに当てなくとも
花が咲くし、生育が出来る。
北半球の太陽は
南に上がる。
南半球の太陽は
北に上がる。
だから、北半球で向日葵、ランも・・・・
花は太陽に向って南向きに咲く。
しかし、赤道から北回帰線の間のエリアでは、
栄養生長最盛期の夏至では、
太陽は北に上がる。
日本人からみれば、夏に太陽が北に
昇るのは不思議であるが・・・。
上の太陽高度の式で計算するとわかる。
カトレアの一枚葉のランは、葉は光を受けるパネル
であるから、当然葉は太陽方向に向く。
北緯の中南米を原産とするランは、夏至には、
北からの光を受けて生育することになる。
パフィオのパービセパラム亜属は、
自生地ではどの方向からの光の場所なのか?
葉の温度と密接に関係する。
反射、散光なら・・・・どうなる?
パフィオと日照・・・散乱日射
この日照には
気象では直達日射と散乱日射がある。
ラン栽培で、この二つの日射が論じられた
ことはない。
不織布による「散光」というのは、
この散乱日射のことである。
今年のような猛暑の時は、直達日射による
紫外線、葉の高温が大問題になる。
気温が高い状態に、更に直達日射の光が
ランの葉に大きなダメージを与える。
自生地の空を、光を日本に当てはめることは
出来ない。
ランにとっては日本は全てが異空間の
異郷の地である。
パフィオは直達日射と散乱日射のどちらを
好むのであろうか?
進化の原点の基本のDNAはどちら?
散乱日射である。
散乱日射というのは晴れた日も、曇りの日も
あまり変化がない。
遮光と散乱日射とは意味が異なる。
1
2
3
3
2
1
Paph malipoenseの葉の反射と集光と吸収を模式化すると
上のようになるかもそれない。
1 葉の表面で反射。
乾期の強光の大部分は反射されるが、雨期の弱い光の緑以外の光は
吸収される。弱光下で緑に見える部分。薄緑青の部分。
2 光は葉の裏の紫まで到達して反射される乱反射光。
弱光下で中間の色に見える光の部分。緑の部分。
3 葉の裏の反射細胞まで透過する間に葉緑素からほとんどの光が
吸収された部分。濃い緑の部分。
緑ということは、緑以外の光が吸収されたということ。
白く見えるということは、全部の光(7色)が反射されたということ。

ラン栽培の基本中の基本は「自生地再現」である。
ここで自生地に行くこと出来ない人にとっては、プラントハンターによる自生地の記録が大変参考になる。
ランの本には栽培環境、条件、管理などは、この自生地の記録に基づいて書かれている。
特に、新しく発見されたランでは、ハンターが発表する記録が唯一の手がかりである。
ここで一番大きな問題は、
プロトコームを発見できないということである。
植物標本、プラントハンターの目的は「成株」であるということ。
成株の写真、状況は相当なところまで観察されるが、
種子からプロトコーム、幼株の姿が見えない。
自生地というのは、必ず、植物では種子が芽生え、生長できる条件があるということである。
ランはラン菌との共生で発芽しなければならない!!
これが絶対の条件である。
大株の自生状況がどんなに過酷に見えても、
種子が発芽出来る条件が備わっているという事実である。
この一点からランの自生地を考察すれば見えてくるものがある。
1 ラン菌がそこに生息していること。
2 ラン菌が生息し、菌糸が伸びる湿度があること。
どんなに乾燥に感じられても・・・・・・
生きられる限界を越えた乾燥のゴビ砂漠、サハラ砂漠には植物は生息できない。
このことからみると、ランの自生地が、どんなに乾燥していると言っても、
ランが生きられる湿度は必ずある場所である。
3 プロトコームがいきつづけ、発育できる湿度が常に保たれていること。
4 生きつづけられる温度があること。
5 葉のあるランでは、生きつづけられる光量、光質があること。
6 大氣があること(当然であるが・・・)空気があるから風も対流もある・・・・。
厳しい乾期のあるところに自生するランの場合、
プラントハンターが自生地に足を入れるのは「乾期」であることから、
日中の環境が記録される強調される場合が多い。
例えば、カトレアのワルケ、ノビリ・・・・・・。
熱帯の乾期。
日中40℃にもなる中での乾期。カラカラ・・・・。
木の葉落葉しているから、灼熱の光が降り注ぐ・・・・。
そういうところでも、実は、プロトコームが生きられる湿度があるということ。
ラン菌が生きられる湿度があるということ。
ところが、我々の栽培では、素焼鉢で水ゴケで植えた場合、
プロトコームが生きられない湿度であることがほとんど。
コルクに付けた場合も同じ。
一口に言えば、ノビリにとって、日本の温室は・・・・・
ブラジルの自生地よりも・・・・酷く、辛い・・・環境ということが出来る。
これが、ラン栽培の現実である。
プロトコームが生きられない乾燥。
ラン菌がいない栽培。
どこかがおかしいと感じませんか?
自生地再現といいながら・・・・・
株は掘って持ってくるが、ラン菌など考えもしないこと。
プロトコームを探さない・・・という研究。
ここの原点からランをみないと、本当のラン栽培は完成しない。
宇井清太はこのように思っている。
SUGOI-neで栽培してみると「おかしい」ことが見えてきたということ。
このことは植物分類学とは次元が異なるエリアのものである。
このことは、パフィオ栽培にも当てはまる。
パフィオ栽培では、分類学で6亜属に分類し、それを基にして栽培法が究明される場合が多い。
しかし、本当にそうか?
6亜属の違いというのは、プロトコームが生きる条件も6つの違いがあるのか。
そういう疑義である。
そこから栽培を考えないといけないのではないか。
プロトコームが生きられる条件に6つの違いなどないのである。
パービセパラムもブラキもポリアンサも・・・。
前記したような共通した条件が、自生地には必ず存在するということ。
雲南にも、キナバルにも、ヒマラヤにも、フィリピンにも・・・。
後は、どんな条件が大きく生育に作用、支配しているかを検索し、
微妙に調節すれば出来るということである。
ランのような新参者は、
進化の基点時代に具備したDNAを持ちつづけているからである。
それが、種子からプロトコーム発生の時、もっとも原始的な姿を見せる。
ほとんど全てのランは、このプロトコーム時代に大きな差、変化はない。
大きな環境適応の変化を見せるのは、プロトコーム以後の生育からである。
別な植物で例を上げれば・・・
ヒマラヤを発生の基点とするシャクナゲ。
南に進出したボルネオ島のキナバル山に自生するシャクナゲ。
ほぼ赤道直下のエリア。
四季はない!!
でも、シャクナゲ、ランの開花期には四季が存在し、開花期が決まっている。
熱帯生まれの熱帯育ちの植物では、一年中花をつけるものがあるが・・・・。
北緯23,4度の北回帰線、南緯23,4度の南回帰線。
これより以北、以南には四季が存在するが、
ヒマラヤを発生の基点にするシャクナゲには、この四季の変化のDNAが存在する。
キナバル島のシャクナゲの開花期。
赤道直下の四季のないところで、なぜ開花期が決まっている?
赤道直下でも太陽の高度差がある。
夏至。
このとき北緯23,4度の地点で、太陽の高度は天頂である。
夏至の赤道での太陽の高度は天頂ではない。
したがって、赤道直下のキナバルでは、それより太陽の高度は低いことになる。
つまり、シャクナゲは、北回帰線と南回帰線の間の夏至、冬至の間で変化する太陽高度で、
季節の変化を感知しているといわれている。
この太陽の周期的変化に合わせて栄養生長、生殖生長が周期的に行われている。
四季の変化のDNAを捨てていないのである。
日本のようにはっきりした四季の変化はなくとも、微妙な太陽の高度差を感知して、
季節の周期性を栄養生長、生殖生長のサイクルにしているのである。
パフィオはどうか?
ボルネオ島のパフィオも開花期は定まっている。
赤道直下に自生するパフィオが一年中不定期に咲くわけではない。
開花期が決まっている!!
そして、赤道直下の島には四季がないにもかかわらず、図鑑では「開花期は夏」と記載される。
植物の世界では、赤道直下の島にも四季はあると認識されいるのである。
だからこそ「開花期は夏」「開花期は春から初夏」と記載される。
このパフィオ定期的な開花期は、生まれは北緯、南緯0度のところで生まれた
植物でないことを示しているのではないか。
一年中、どれかの株が咲いているようには進化していない。
不定期咲き・・・・でない。
キナバルのパフィオもシャクナゲと同じように四季のDNAを捨ててはいない。
このことはキナバルに自生するCymbidiumにも言えることである。
おおくのCymbidiumの原種が自生するが、
開花期はほぼ定まっている。
この生長周期は雨期、乾期の単純な周期ではなく、四季の変化の周期である。
赤道直下の島・・・
なのに!!
そういう意味で、進化のもっとも基点になる条件のところから、
栽培条件を検索すれば、見えてくるものがあるのではないかというのが、
宇井清太の考えである。
パフィオもCymbidiumもヒマラヤを進化の基点とする植物である。
両者には栽培上の共通性を見ることが出来る。
ナドソンは100年前に、ランの進化の基点となる炭素循環の糖を発見した。
これがランの栄養の基点だから、現在でも、この技術は廃れることなく使われている。
他のことはアレコレ変化しても、発芽、苗生産では、絶対の技術である。
不滅の基点である。
NEXT パフィオとラン菌
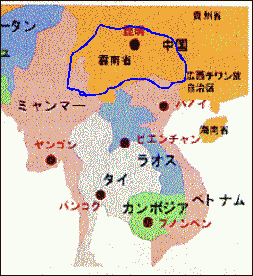
上の地図の青色のところはおおむねではあるが
東洋蘭の「中国奥地蘭」の自生地である。
この自生地とパフィオのパービセパルム亜属の自生地
は重なる。
つまり同じ気候の下で生き続けて来た。
ということは、中国奥地蘭の栽培環境と、
パービセパルム亜属の栽培環境には
共通点があると考えることが出来る。
非常に短絡的な見方にも見えるが、
こういうことが多にして、的を射ている場合がある。
中国奥地蘭はCymbidiumなので、
SUGOI-neで素晴らしい生育をする。
パービセパルム亜属は原始的か?
自生地の特異なライムストーンの反射光の下で、
集光レンズを具備した姿を見ると、
原始の姿より、より進化した姿とも見える。
集光と反射は、自生地の特異な光条件と
モンスーン気候の変化が複雑に絡み合った
気象条件、気候サイクルに適合した姿であるからである。
ライムストーンの不毛の場所では、
一般の植物は生きることが出来ないからである。
同じ場所で他のランは生きられないからである。
例えば、カリマンタン島には、標高の違いがあるが、
同じ場所に多くの植物が、ランが自生できる条件がある。
島全体では1200種に及ぶランが自生している。
その形態は多様であるが・・・・
それらのランには共通した環境へのDNAがあるからであ。
集めた光を有効にするには・・・
葉を薄くする必要がある。
「陰葉」である。
葉の厚い原種でも、日除けを強くすると、
葉を薄くする。光を有効に使うためである。
エビネ、リカステなどは典型的な陰葉。
集光レンズ細胞で集められた光は、
葉肉を通過して反射する。
一般の植物の光沢ある葉では、
葉の表面のクチクラ層で反射させる。
同じ反射でも全く異なる光利用である。
Paph malipoenseの自生地。
そこはライムストーンの反射光の世界である。
喬木からの木漏れ日の光の世界ではない。
雨期にには、このライムストーンの反射光に、
更に霧、曇天の散光が加わる。
つまり葉の表面を覆う集光レンズ細胞は、
雨期には光を集め、乾期に強い反射光の時は、
葉の裏にある紫の反射細胞によって、
余分の光は放射するようにしている。
相反する光条件を見事にクリアしている。
不毛の地に生きる新参者の智恵である。
同じ自生地には他の多くの植物も自生する。
しかし、それらの植物は条件の良いところを
占有している。だからPaph malipoenseのように、
ギリギリの極限に近い葉の構造を備える必要はない。
同じエリアに自生するCymbidiumの
いわゆる「中国奥地蘭」には、集光レンズ細胞はない。
光に恵まれた場所に自生するランには
葉肉の厚いものが多い。
気孔からの炭酸ガスが内部の細胞に
届きにくいが、そのマイナスと、水分
貯蔵の機能のプラスと、微妙な計算が
あっての進化であろう。
カリマンタン島の葉肉の厚いパフィオの原種と
ファレノの原種の葉肉の厚さには共通点が
みられる。
葉肉の薄い葉は、
光は葉肉の細胞に光が届きやすい。
したがって、葉の薄いランは、弱い光を
有効に利用することが出来る。
コチョウランの原種にも、
同じような葉の模様と、葉の裏が紫のものがある。
Pha
celebensis, lindenii, lowii, philippinensis,
schilleriana,
光条件に共通性をみることが出来る。
裏紅の植物には他に
ウラベニショウ、ウラベニシュウカイドウ、
ウラベニ大文字草などがあるが、
ウラベニ大文字草はPaph armeniacumと同じように
石灰岩の反射光のところに自生する。
研究すると面白い共通点があるかもしれない。
カラテア マコヤーナ の葉の裏
この葉の裏の紫は、Paph armeniacum
などのパフィオを極めている。
透明に近い細胞もほとんど同じ。
カラテア マコヤーナ
ブラジルの熱帯雨林の下草として自生する。
黄緑のところは「透明細胞」である。
この部分に当たった光は、細胞を透過して、
下の葉に当たる。
つまり、光を下の葉に分けて、全部の葉で
光合成を行なえるようになっている。
楕円形の巾広い葉でも、これなら、下の葉に
濃い影を作ることはナイ。
このような光のことを考えると、
パフィオの場合、葉がピンとたって、
一枚目と3枚目の葉の間に、大きな間隙
があるような状態でなければならないことになる。
この間隙に横からの散光が入り当たることになる。
葉が垂れるような状態では、充分な光合成が
出来ないことになる。


カランテ(エビネ)の葉も、パフィオよりは細胞が小さく緻密であるが、
パフィオと同じような集光レンズと反射鏡のクチクラ層を持っている。
エビネの葉の光沢は、この細胞の乱反射によるものである。
照葉樹のアオキ、ヤツデ、椿。
ランのデンドロ、コチョウラン、マスデ。
このような照葉は、パフィオ、えびねの葉の光沢とは根本から異なっており、
集光レンズも反射鏡の構造ではない。
パフィオ、エビネのような葉の細胞を持ったランは、
上からの光でなく、散光の光を無駄なく集めるように進化したと考えられる。
このことから、パフィオ、エビネ栽培では、ダイオネットのような隙間のあるものでなく、
霧の光のような散光が適していると考えられる。
実際に散光の不織布の下でパフィオ、エビネを栽培してみると、
素晴らしい生育で、夏負けしないことがわかった。
このことから、ランの遮光について考える場合、
拡大鏡で葉の細胞を調べ、集光レンズを持つランと、持たないランを分類し、
それぞれにあった光の下で栽培すると良いのではないか。
昔、イギリスで、パフィオの温室に石灰乳を塗って「散光」にしたが、
まことに理に適ったことである。
英国の高緯度の太陽の光でも、そのように光対策を行った。
光については、葉の温度も上昇させ、この輻射熱は、
葉の周囲の微気象をも変化させ、葉の周囲の空気を乾燥させるので、
相当深く再研究しなければならない。
葉の温度上昇は、更にハダニの発生とナンプ病を誘発させるからである。
ランの光。
一般の植物と同じように考えてはならない。
右の写真はブラジルの熱帯雨林に自生するCALATEA MAKOYANA.
喬木の下にコボレ日を拾って自生する。
現在は自生地の場所がブラジルとアジアの違いはあるが、
何千万年前まで遡れば、中南米、アジアなどの分け方はないから、
新参者のパフィオが喬木のコボレ日で生きる進化の道を選んだ時、
先輩植物をモデルとして進化したと考えられる。
したがってPaph armeniacumなどの葉裏が紫のパフィオを栽培する場合、
光条件はカラテアの光条件を参考にすれば良いともいえる。
園芸では「明るい日陰」と表現する。
これは「散光」である。
クチクラ層がレンズになった大きな細胞。
強い光では、この光を集めるレンズが仇となって「葉焼け」する。
光阻害である。
このカラテアは、光に敏感な反応し、光が強く乾燥すると、
極端に反応し「ヘタッ」となる。
光が散光であれば、葉は「ピン」と立つ!!
葉の「ピン」・・・・でベストな光がどうか判定できる。
葉裏の紫が全面にないところが・・・・自生地における光条件と
深く関係し、絶妙な集光と反射と透過光で光合成を行なっている。
「光阻害」について
ラン科植物の中にも・・・「明るい日陰」・・・散光でなければならないものが多くあり、
強い光は「光阻害」となって大きなダメージを与える。
光はある一定のところまでは光は強いほど光合成速度は増してゆく。
ある強さ以上になると、逆に光合成は減少する。
これを「光阻害」という。
植物は、アレヤコレヤの対策を講じて光阻害の備える。
前記の写真の透過、反射もその一つである。
葉の角度もその一つである。
垂直に立てれば、真上から直光は・・・葉に当たらない!!
それでも光合成に必要以上の光のエネルギーは何処にゆくのか?
光は光合成に必要であるが、強い光の場合は、それ以上の光合成は出来ないから、
この余分のエネルギーは光合成に使うことは出来ない。
この場合、前記したように植物には防御システムが備えてあり、
ある程度まではエネルギーを安全な熱エネルギーに形にする。
この熱が細胞温度を上昇させ光合成に関係する酵素(たんぱく質)の機能を低下させる。
しかし、更に強くなると、余ったエネルギーは「活性酸素」を作り、
光合成システムを破壊する。
紫外線などによる「活性酸素」が光阻害の原因ということが近年解かってきた。
宇井清太が、このホームページでしつこく「紫外線カット」というのは、
このことである。
ハダニに大発生。
これが光阻害である。
一つの条件が、一つで終らないで、多方面に関与するところに、
生き物の奥の深さがあり、栽培の難しさがある。
Cymbidiumは「光を好む」と無造作に書いてある本が多くあるが、
本当にそうか?
光の何を好んでいるのだ?
光と一口に言っても、光の種類、質から・・・・いろいろ。
だから、ランは本を読んだ人ほど作れないということが出てくる。
パフィオ栽培では、市松模様、ダイオネットでは、隙間から、直光が差し込むが、
この光が強いと光阻害となって、株を大きく弱くする。
当然ナンプ病の原因となる。
この光阻害を防止するのが不織布の散光である。
この散乱した光を集める細胞をパフィオは具備した。
クチクラ層の機能を減少させてである。
この進化を無視してはならない。
自生地において、常緑のパフィオの葉に当たる光は、
一日の中にも、一年の中にも大きな変化がある。
少ない時は「集光」。
足りない時は「反射」と「透過」。
これで光を調節する。
葉の角度、葉と葉の間隔、葉の面積・・・・。
微妙に変えることまで行う。
光阻害は・・・・
低温、乾燥などのストレス条件で強い光を当てると、比較的弱い光でも光阻害を受ける。
パフィオ栽培で、石油高騰で低温栽培の場合は、弱い光で栽培しなければならない。
一番間違いが起きやすい場面である。
低温栽培では、根腐れがコワイから乾燥させて作る。
乾燥と、強い光は・・・光阻害を大きくする。
SUGOI-ne栽培では、適温より低温で作る場合は、
より弱い光で絶対にオシボリより乾燥させてはならない。
カトレアも、高温性のパフィオもカラカラ、強光では、極端にかぶが弱るが、
カトレアは耐えて耐えて我慢しているのに、
人間は・・・・好むと理解してしまう。
なぜ多くのランの葉は緑色なのか?
緑色に見えるというのは、光の中の他のいろが吸収されて、
緑色が吸収されなかったということ。
だから、森の中には緑の光が一杯ある。
熱帯雨林の光の弱いところには「紫色」の植物がある。
ランにもある。
こういう場合は、喬木がこぼした緑の光を拾って光合成している。
したがってPaph malipoenseの葉の裏の紫の細胞は、
光の反射の為の細胞であると同時に、喬木がこぼした緑の光で、
光合成していることになる。
葉の表面の集光レンズ細胞は、このために具備されたと考えられる。
考察
パフィ栽培での謎は・・・
パービセパルム亜属がもっともなぞめいている。
しかし、ラン科植物の基本中の基本は
1 菌根菌植物であること。
2 自生地にはプロトコームが生きられる条件があること。
3 自生地では新参者植物であること。
この三つの方向から、他の植物との類似性を検索しながら詰めてゆけば、
謎が・・・解けてゆく・・・・。
パフィオのレンズ細胞は、カランテの葉にもみることが出来る。
そうすると、光の類似性が見えることになる。
不織布を張ると、パフィオもエビネも嘘のように素晴らしい生育をする。
ダイオネット、それの二重張りでは不織布の光あ創れない。
霧の乱光・・・散光・・・影の出来ない光。
葉の集光レンズへの進化が見えてくる!!
紫外線カット、散光・・・・葉の温度検温による潅水、換気。
ペクチン潅水法でほとんどパフィオ栽培法は完成しているのではないか?
SUGOI-neのラン菌。
パフィオの亜属による変化は・・・どう考えても・・・・そんなに大きいとは考えにくい。
それより、温室栽培の場合は、葉の温度の検温をしたことあるのか、ないのか。
大きなポイントである。
Cymbidium栽培では、ここがポイント。
室温より葉の温度を高くなるような管理では、ほとんど失敗する。
温室より「蘭舎」の方が合うかもしれない。
山野草栽培の感覚。
Cymbidiumの春出しの切花栽培出来る温度管理と同じかもしれない。
高温性のパフィオとは、まるで温度管理が異なっている。
これは、Cymbidiumにもいえることで、
鉢物栽培と切花栽培では、まるで栽培技術は異なる。
現在、Cymbidiumの鉢物栽培している人は、
ほとんど春出しの切花栽培は出来ないかもしれない。
切花栽培で経営できる技術を持つ人は、
鉢物栽培は簡単に出来る。
ラン栽培というのは、そういうものである。
栽培というのと分類学が相当異なる場面が出てくる。
パフィオでは6亜属13節に分類しているようであるが、
Cymbidiumの変異は膨大で、分類学では手に負えない世界をひろげている。
花による分類では限界があるという一例である。
逆にいえば、分類できる種というのは領域が狭いということである。
栽培に必要な栽培技術の巾が狭いということも出来る。
例えば、Cymbidiumでは、ヒマラヤを基点として北海道の利尻島まで北上する。
この距離の中に膨大な変異株を作りながら、
雪に埋もれても生きられる遺伝子を作り上げた。
つまり、パフィオは、半年も暗黒と低温の雪に埋もれてまで生き続ける進化はしなかった。
シャクナゲとCymbidiumとパフィオは非常に似た生息エリア拡大、進出の仕方をした植物であるが、
パフィオは低温、雪、暗黒という条件を克服しなかったランである。
ツツジ科植物とラン科植物は、過酷な条件を菌根菌との共生でしのいだ。
強酸性、石灰岩・・・この両極端なPH条件。
SUGOI-neは強酸性のツツジ科植物も素晴らしい生育をする。
石灰岩に自生するパフィオも素晴らしい生育をする。
これは何を意味する?
菌根菌の働きを意味しているのではないか?
富士山にも登山ルートが何本もあるが・・・・・
別なルートから登らないと、全体像は見えない。
ひょっとして・・・・別なルートに・・・本当の姿が見える場合もある。
人間を理解するのに・・・オランウータンを観察すると・・・・見えるものもある?
そういうことかもしれない。
クラブに所属すると、ある程度までは情報を得られるが、
その空気が、レベルが固定観念、組織拘束となって、自由な発想を邪魔にする。
そういう場合がむしろ多いのかもしれない。
ランの栽培では、ここのところが難しい。
本を多く読んだ人ほど上手く作れない場合が出てくる。
この例が・・・根のペクチン。
花の審査では、モノスゴイ鑑識眼を持っていながら、根のペクチンを観察しない。
こういうことが出てくる。
木を見て森を見ない。
そういうことが、SUGOI-neで解かってきた。
常に森を見ている人は、SUGOI-neで無造作に素晴らしい生育になった。
細かいところアレコレ見ている人は・・・SUGOI-neを使いこなせない。
権威者なのか、権威者でないのか。
SUGOI-neは検定する試薬みたいなもの。
山の草栽培に、権威など要らないのに・・・・。
コチョウランの葉の表面。
左のパフィオの写真と同じ倍率で撮ったものであるが、
コチョウランの葉の表皮には、パフィオに見られる
細胞はない。
ほとんどの植物は、このようなクチクラ層を持っている。
光の質が葉の進化に密接に関係している。
Paph ツヤ イケダ ’オオイソ’
光沢ある葉を持っているが、この葉の光っているところを
拡大してみると、葉一面が反射鏡のようになっている。
この細胞は、集光レンズと反射鏡の両方を行っていると考えられる。
この反射光で光沢あるように見える。
ツヤ イケダの葉の裏は紫ではない。
葉肉は光を透過する。
ここのところが同じパフィオであってもPaph armeniacumなどのブラキと異なる。
パフィオの葉の表皮細胞は、このような構造になっている。
光は植物にとってもっとも大切なものであるが、
強すぎる光は、逆に植物にとって害を及ぼす。
太陽の光は、植物に都合よく日の出から日没まで
照射しているわけではない。
陰生の植物であるランにとっては、最適な光などは
滅多にないのかもしれない。
弱すぎる。
強すぎる。
こういうことであろう。
強い光は葉の温度を上昇させる。
この温度を冷やす、逃がす方法が出来て、
陸上で生きられるようになった。
同時に必要以上の光を逃がす方法も
行われている。
この限界を越えたとき、植物は生きてゆけない。
あれもこれも総動員して、葉を光から、高温から
守っている。
ランの光利用の効率の悪さ。
太陽からの同じ光を受けながら、
トウモロコシのように、
100日位で2mも伸びるものもあれば、
パフィオのように2,3mmというものもある。
人間にはモット、MORE・・・という向上心が
あるから、2,3mmの生育には我慢できない。
モット・・・・早く、大きく・・・と想う。
あげく、肥料で、技術で何とかすると考える。
ラン菌の居ないコンポストで植えていながら、
こういう無理難題をランに押し付ける。
果樹栽培でも、
桃、ブドウ・・・などでは、植えて1~3年の間に、
窒素肥料を多く与えれば、早く生長はするが、
冬に凍み枯れ、眠り病、胴枯れ・・・
非常な短命になる。
自然界の炭素循環、窒素循環以上には、
若木には与えてならない。
パフィオ栽培者の多くは、
農業を知らない。
作物栽培と、原種栽培は似ているが異なる。
現在のランの肥料は、
作物栽培から移行して発想したものである。
厳密な意味での「菌根植物」であるランの
養分吸収 システムの肥料?とは根本から異なる。
正確にいえば、ラン栽培では「肥料」と呼称
すべきでないのかもしれない。
光合成のエネルギーのみで生育可能な植物と
ランを同じに考えれば、野菜に使う肥料でも良いことになるが、
ランというのは炭素循環の「糖」をエネルギーにするので、
即、同じで良いということにはならない。
SUGOI-neの開発で、
ここのランの根本のところが、ようやく見えてきた。
ラン栽培は今後大きく変わる。!!
反射される光。
必要以上の光は反射細胞により、
葉の外に放射される。
陽生植物から見れば、まことにもったいないこと。
「足りるを知る」!!
パフィオのパービセパラム亜属、ブラキペタラム亜属は、
この精神で生き続けているのか?
森の不毛の場所で分を守り・・・
つつましく生きるのか?
集光細胞の謎
なぜ?
パフィオのパービセパラム亜属、ブラキペタラム亜属は、
「集光」と「反射」の相反する機能を持つ組織を具備したのか。
葉の表面の反射でなく、葉肉を透過させた光を反射させている。
しかも、他の種より葉肉は薄い。
ここに光に対する進化の謎がある。
光を集めるということは熱をも集めるということ。
葉肉は、この熱と深い関係がある。
葉肉を厚くすれば「蓄熱」「保温」することになる。
必要以上な高温を防ぐには・・・・蒸散、気化熱による冷却が必然になる。
水の乏しい場合は、このシステムでは致命的な欠陥になる。
低温時期の光合成の熱は・・・どうやって調達する?
・・・・・。
進化の謎が見えてくる。
見えれば、栽培は容易になる。
しかし、植物の進化、生存というのは不思議である
陰生植物である、ヤツデ、アオキ、クサソテツ、タニワタリ・・・などの葉も、
光沢をもって光を反射している。
パフィオ、フラグの葉と同じ機能を持つ。
デンドロ、ノビル系の葉もピカピカの光沢。
エビネには光沢のあるもの、少ないもの・・・・。
この葉の光沢・・・反射は・・・・?
紫外線?
余分な光は・・・・葉に取り込まないで・・・返品?
パフィオを栽培する場合、この葉の光沢のあるもの、ないもの、少ないものを、
この意味を考えて光対策をしなければならないのかもしれない。
皆さん、研究してください。
光合成の効率から見ると
左の写真は、モンスーン気候の下で進化した
Cymbidiumとパフィオ。
同じような光条件、雨で光合成を行ないながら、
この差は・・・どういう意味を持つ。
Cymbidiumのエネルギー蓄積は偉大である。
このCymbidiumのバルブの重量は700gある!!
10個のバルブの株なら7kgである!!
この株が10000株なら・・・・・70000kg。
実に70t。
これをわずかな肥料と光と水で作り蓄える。
それと比較するとパフィオは・・・・・となる。
エネルギーを蓄えることを放棄した?
光の吸収。
光の反射。
パフィオのパービセパラム亜属、 ブラキペタラム亜属などは、
「向上心」のDNAを一部放棄したのではないか?
ささやかに・・・清く・・・
でも・・・雨にも負けず・・・・風にも負けず?
しかし、光の有効利用、効率から考えると、Cymbidiumとパフィオでは、
エネルギーの蓄積が比較するのもどうかというほど、その差は大きい。
同じランでありながら、進化の方向が、ここまで異なるのである。
この光エネルギーの蓄積とパービセパラム亜属、 ブラキペタラム亜属の
レンズの細胞、葉の裏に備えた反射細胞は、深い深い関係があると宇井清太は想う。
光の反射。
光の放出!!
普通なら・・・・放出するということは・・・もったいない話である!!
考えてみると、ランの愛好というのは、
不思議な人間の感覚である。
こんなに生育の遅い、効率の悪いものを・・・・イイという。
こういう遺伝子を・・・・。
なぜ大切な光を反射放出する?
なぜ身体を小さくする進化を選んだ?
パービセパラム亜属、ブラキペタラム亜属に見られるこの生き方は、
ラン菌との共生と深い関係があると考えられる。
つまり、ランには葉による光合成を放棄した腐生ランが存在する。
ラン菌が炭素循環でもたらす糖のみで生存しているランがある。
ここまでランというの植物は狡猾な生き物である。
このことに着目すれば、パフィオの光反射も、極端に生育が遅いのも、
身体が小さいのも説明がつく。
植物が生きる最終目標はは花を咲かせ種子を実らせ、子孫を繁殖させることである。
この目的が達成できれば、他のことは枝葉末節である。
何も大きい葉を持つ必要はない。
何も大きい身体を持つ必要はない。
大きいということはエネルギーを多く必要とする。
スモールは良いこと!!
森の支配者は樹木である。
それと同じ道を歩む必要はない!!
大きい会社は、大きい収益を上げなければならない。
巨大会社、大きい銀行が・・・もろくも消えた!!
世界の巨大蘭園も相次いで消滅した!!
パービセパラム亜属、ブラキペタラム亜属がパフィオの進化で、
「原始」なのか?
それとも最も進んでいるのか?
腐生ランというのは原始の姿なのか進化した姿なのか?
パービセパラム亜属、ブラキペタラム亜属のパフィオが、
腐生ランに近い栄養系の進化であると宇井清太は考えている。
そのように考えると、集光レンズ細胞、反射細胞のほとんど説明がつく。
そうであるならばパービセパラム亜属、ブラキペタラム亜属のみならず、
他のパフィオにおいても、ラン菌との共生による炭素循環の糖が、
非常に重要になるということになる。
ブラキペタラム亜属がパフィオの中で「ナンプ病」に弱いとされているが、
自生地において今日まで生き続けてきたということは、
特に弱い遺伝子を持った種ということはありえない。
自然界において、そのような病害菌に弱い遺伝子を持った種というのは、
自然淘汰の法則で絶滅するからである。
自生地から「拉致」して、ラン菌削除のコンポストで植えている栽培上の欠陥こそ、
最も重要な問題である。
なぜなら、前記したように、光の反射細胞を具備したことは、
光による光合成の糖を、より多く製造するということを放棄した種と考えられるからである。
ラン菌がもたらす糖があるから・・・・間に合っている。
そういうことと考えると、集光レンズ細胞、光反射細胞という相反する細胞を具備した
説明がつくからである。
ナンプ病に弱いのは、我々がラン菌削除のコンポストで植えるからである。
自生地にある炭素循環の糖が鉢内にないからである。
肥料を与えた場合、当然硝酸態窒素の過剰吸収が行われる。
自生地にない高濃度の硝酸態窒素。
ナンプ菌は硝酸態窒素が大好き。
森の中に置けばいいものを。
そんな気持ちになってくる。
7年6月18日
7年8月2日 木曜日 19:03:34
約40日でこの程度の生長。こんなに遅い生育の植物が生きられる空間というのは、
他の植物が生きられない不毛の場所である。農業的見地から見れば、最低の光合成効率となる。
作物という視点に立てば、使い物にならない遺伝子。
この生育の遅さが、市場ビジネスにはならないが、愛好という限られた分野では珍重される。
人間というのは面白い感覚の持ち主である。現代の時間の流れの速さと逆行した進化。
こんな小さな株、あまりに生育の遅い株が、
生き残り生長するには、光合成に深い仕掛けがあるに違いない。
自生地の気温のデーターを鵜呑みにしてはならない。
集光レンズを持った意味は何か。
どんな温度のどんな光のとき、そのレンズは働くのか。
集めた光は、葉の温度上昇にどのように作用しているのか。
自生地の葉の最低温度から・・・どのように温度上昇しているのか。
これを温室に再現しないと、このレンズ細胞が集めた光が、
逆に生育阻害要因になる場合も出てくる。
ナンプ病。
写真のPaph armeniacumは
SUGOI-ne2号ゴールド植え、紫外線カットフィルム、
不織布による散光条件下で栽培。
至極順調。
Paph armeniacum
上の写真は「ユキノシタ」。
Paph malipoenseの葉と極似する。
葉の表面を拡大すると、Paph malipoenseと全く同じ集光レンズの細胞である。
葉の裏も全く同じ。
光条件に対する葉の構造、機能には類似点が多くあり、
パフィオ栽培では、パフィオのみを研究するのではなく、
他の植物から類似点を探し、諸条件を追及することが、
成功への近道かも知れない。
ユキノシタも相当弱い光から、直光が当たる場所まで生育可能である。
しかし、基本は・・・レンズの細胞という点から考えると、
弱い光の下で栽培することが基本である。
しかも集光という機能からすれば、このような葉を持った原種では、
「散光」が基本である。
他の植物に見る集光器を持った葉。
パフィオの葉を見てきたが、光を集める、透過するレンズを持ったクチクラ層は、
他の植物でも見ることが出来る。
自生地における光条件に非常に似たところがあり、
新参者のランは、それらの先達植物の進化を真似たと考えることが出来る。
このように考えると、ランが全く別に進化したのではなく、
植物進化の多様な中から、都合の良い部分を取り入れて環境に適合するように進化したと考えられる。
光沢のある葉と光沢のない葉
パフィオには光沢のある葉を持つ種と、パービセパラム亜属、 ブラキペタラム亜属にみられるように、
葉に光沢のない葉の種がある。
図からも解かるように、光沢のない葉は光をクチクラ層で反射しないで、葉の裏の紫の細胞まで
光が到達した後に一部反射する。
光沢のある葉は、光の多くを葉の表面のクチクラ層で反射する。
したがって、このような葉を持つパフィオは、葉で受けた光の一部を光合成に利用する。
光を当てて段々強くしてゆくと、それにしたがって光合成は増大する。
しかし、あるところまで達すると、それ以上強い光を当てても光合成は増大しない。
ここのところが植物によって大きく異なり、パフィオの原種においても、
種によって大きな違いがある。
遮光を考える場合、この照度の違いと、葉にレンズを持っている種か、持たない種かで、
光の質を替えなければならない。
散光を好むもの。
弱い直光を好むもの。
光合成の補償点
光合成と呼吸作用がプラス、マイナス・・・ゼロになるところ。
植物は光合成を行うが、同時にエネルギー消費の呼吸作用も行っている。
見かけ上、合成と消費が同じになる場面がある。
ラン栽培で最も注意を要する光と温度の問題である。
拡大 太陽電池のパネルのような構造をもっている。
葉一枚に何千何億の集光細胞を配置している。
光は充分あれば、こんな装備をしなくていいはずである!!
陽生植物には、このような葉を持つものはナイ。
このレンズのような細胞の表面は平ではなく凸レンズ又は球である。
ビー球。
戦後間もないころ、道路標識はビー球を集積して作製していた。
それと全く同じ。
集光と反射、乱反射。
パフィオの原種中ではパーセパルム亜族の珠がもっとも大きい。
パフィオの起源、進化から栽培法を進める場合、
この集光レンズ細胞による分類は、相当的を得ているかもしれない。
光の基本がわかれば、その他の条件はSUGOI-ne栽培は簡単だからである。
葉肉を透過した光
葉の裏が紫の葉に比べ多くの光を透過している
透過した光
クチクラ層(葉の裏)
海綿状組織
柵状組織 光合成を多く行うため、細長い細胞を持つ植物が多い。
クチクラ層 葉緑素を含まない細胞
反射した光
反射した光
葉肉の薄い葉
葉肉の厚い葉
クチクラ層(葉の裏)
海綿状組織
柵状組織 光合成を多く行うため、細長い細胞を持つ植物が多い。
クチクラ層 葉緑素を含まない細胞 。
透過した光
直光
反射によって葉の外に放射した光
上の写真のように葉は光を多く透過する。
比較的多くの光を受けるところに自生する種は、このように余分な光は透過させることによって、
光を調整している。
このような葉には表皮に紫外線から守る「クチクラ層」を備えているから、
反射による光の調節ではなく、透過による調節になる。
気化熱による葉の温度調節にはCAM植物は限界があるので、
この光の透過は重要な意味を持つ。
緑の葉を持つパフィオには
葉肉の厚いもの。
葉肉の薄いもの。
この二つがあるが、当然、それには自生地における光条件が関与している。
緑の葉を持ったパフィオはどうか。
葉の裏の紫を拡大して見ると、球状の細胞が重なり表面を覆う。
ガラスの鏡の裏を見ると、同じようにこのような皮膜になっている。
入ってくる光は透過することなく反射する。
ダイアモンドはカットの仕方で裏に皮膜をしないでも100%反射するようにした。
ダイヤの輝きとパフィオの葉の細胞・・・・。
同じ光の反射である。
光の反射は女性の心を捉えると同時に、パフィオを生存させるものでもある。
散光、弱光下の葉の光の反射、透過。
直光下の葉における光の反射、透過。
直光は多くの光が細胞を透過する
光不透過細胞
光を反射させる
ほとんどの光は反射する。
葉緑素を持った細胞
光合成細胞 この細胞を光が通過するとき
光のエネルギーは利用される。
集光レンズ細胞
上の図
光の強い場所での光の反射である。
光が強い場合は、葉の裏の黒紫の球状細胞に光るが当たると、
光は反射し、この光は葉の透明細胞を通過してゆく・・・。
葉肉は薄く光合成細胞は少ないので、光の余分なものは吸収されることなく、
葉の外に出てゆくことになる。
光の中には赤外線、遠赤外線のような電磁波も含んでいる。
葉の温度上昇の対策は、葉肉を薄くして蓄熱しない構造にしなければならない。
葉が薄いということは光合成出来る葉緑素が少ないこと。
パービセパラム亜属、ブラキペタラム亜属にみることが出来るが、
最小限のエネルギーで生存する道を選んだ種といえる。
その代わり不毛の地でも生きられる適応性を持ったと考えられる。
光というのは葉のある植物にとって必要なものであるが、
余分なもの、必要以上に強い光は、逆に害を及ぼす。
この対策に植物は長い時間をかけなければならなかった!!
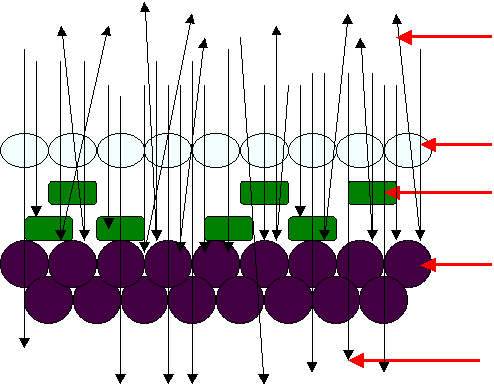
写真1のPaph malipoenseの葉の集光、反射、透過は上の図のようになる。
透明の表皮細胞(集光レンズ細胞)を通過した光は、光合成細胞の隙間を通って葉の裏の
光不透過細胞に当たる。一部は透過するが大部分は鏡に当てた光のように反射する。
反射した光は光合成細胞に当たり、光合成のエネルギーとして利用される。
この光を全方向に反射させるには全方向から入射する光でなけれなばならない。
散光。
霧の中の光である。
霧は水の粒子であるから、太陽からの光はこの粒子に当たって「乱反射」する。
この乱反射した光をPaph malipoenseのは最も効率よく集める機能を具備している。
このために、葉を幅広く大きくする必要はなくなった。
葉は受光パネル。
この機能を持たないエビネ、リカステなどは、身体に不釣合いな巨大な葉を持っている。
同じ散光を集めるのであるが、進化の方向が異なれば、このような葉の構造の違いになる。
林床の光の乏しいところに地生するランにとって、
いかに多くの光を集めるかが進化の最大のポイントになる。
必要は発明の母である。
光のサンサンと降り注ぐ場所に自生する植物の葉は、光を集めることよりも、
紫外線から身を守る、高温から身を守る進化である。
パフィオの自生地は種によっては、極端に光条件の異なる場所でも生息している。
どちらが本当の光条件なのか解からなくなる場合が出てくる。
適応性の巾・・・と呼んでいるが、この葉の細胞を顕微鏡で観察すれば、
容易に散光がDNAに組み込まれているものが一目で判明できる。
なぜ光の適応性の巾が広いのか?
このような集光細胞を持ったパフィオはパービセパラム亜属、
ブラキペタラム亜属にみることが出来るが、
その種の中には林床の光の弱いところから、直射光線まで自生できる機能を持つものがある。
他のランでは、このように光の幅は広くはないのが普通である。
では、なぜ・・・このような幅広い適応性を身につけたのか。
上の図は光の弱い条件の場合のである。
この場合の反射した光は弱いから光合成細胞でほとんど吸収される。
光の強い場合はどうか。
非常に功妙に光合成細胞が配置されている。
葉に斑点があるのは光合成細胞(葉緑素のある細胞)と
葉緑素のない透明な細胞が斑に配置されているからである。
この斑模様は他の植物の突然変異の斑入り植物とは根本から異なる。
そういう細胞の配置による模様は伊達や酔狂でしているのではなく、
光環境に適応するために進化して、そのDNAは確定して次世代に継がれてきたものである。
新参者のパフィオが生きる場所は限られている。
しかし、種族の繁栄は生物である限り絶対のDNAである。
ならば・・・・林床から抜けて・・・海岸の岩壁、石灰岩・・・・
陽光の直射する不毛の地をも開拓して生息しなければならない。
弱い光のところでも、強い光のところでも光合成出来る葉の機能を持つようにする。
このように進化する方向は当然ありえる。
透過した光
透過する光は非常に少ない。
光不透過細胞
光を反射させる
ほとんどの光は反射する。
葉緑素を持った細胞
光合成細胞
反射した光は、この細胞に当たる。
集光レンズ細胞
光の透過
左の写真1,2は同じ条件で光を照射して、
葉の裏からみた葉の明るさ。
写真1はPaph malipoenseの葉。
鏡の裏のようになっているため、光は葉裏を透過する量が少ない。
写真2は ツヤ イケダ の葉。
光は葉肉を透過している。葉で受けた光の利用が少ない。
光合成には効率が悪いが、光を通すことは、光の熱線をも透過する。
比較的光の多い場所に自生するランは、ほとんど、このような
光の透過である。
葉の裏に光を通さない黒紫の細胞を配置した。
この自然界の妙趣というか、進化の奥の深さ・・・・
ガラスの鏡。
ダイヤモンドのカット。
道路標識。
光の透過と屈折と反射・・・・人間が考える前に、
パフィオは身につけていた!!
黒紫の細胞に集められ到達した光は、どのように葉緑素を持った
細胞にあたり、光合成を行なうのか?
透明な細胞で無駄なく集められた光。
それが黒紫の無数のまるい細胞に当たると
あらゆる方向に反射することになる。
葉肉内の細胞に光線は当たることになる。
この光の当たる量は、弱い光であっても
陽光植物の一般の葉の利用する光の数倍にも達するといわれる。
Paph malipoense
写真2
写真1
パフィオのウラワザ。
多くの光を集める究極の機構と機能
Paph malipoenseの葉で見てみよう。
1 葉の表面に無数の集光器のレンズを装備した。
写真参照 約30倍拡大。
トンボの複眼のような感じ。
散光の光を効率よく集めることが出来る。
2 葉の裏に濃い紫の球の細胞を装備した。
鏡の裏を黒くしているのと同じ、この紫の細胞で光の透過を遮断し、
球状の玉は乱反射し光は葉肉の葉緑素に多くの光が照射するようになる。
ランは森では新参者。
植物にとって良い環境とは最高に光合成出来る場所である。
しかし、ランに残された場所は、他の植物が見捨てた場所である。
その中で光条件はランにとって最大の悩みである。
喬木がこぼす光。
その弱く少ない光では充分な光合成は出来ない。
林床の光の少ない場所に自生するランは光を集めるのに工夫する。
エビネ、リカステ・・・などは葉を薄くして幅広くする。
パフィオ パービセパラム亜属、ブラキペタラム亜属は、
逆に葉を小さくしてエネルギーの消費を少なくして、
多くの光を集める究極の機構と機能を備える進化をした。
著作権所有者 宇井清太
引用、複製、転載など著作権に関わること全て
禁止いたします。
反射鏡と
集光レンズを持つ・・・パフィオ
進化の謎に迫る 宇井清太
konnposuto 1054